Start learning
The development of the digestive system involves the formation and differentiation of various structures, including the esophagus, stomach, liver, gallbladder, pancreas, and intestines.
Development begins with the emergence of an epithelial tube enveloped by splanchnic mesoderm. The mucosa undergoes rapid development, leading to the formation of folds, while the mesenchymal covering differentiates into various layers.
The digestive tube is divided into different segments, each with specific characteristics and functions. Mesenteries, double layers of peritoneum, anchor segments of the digestive tube to the trunk walls and allow for the passage of blood vessels and nerves.
The foregut gives rise to the esophagus, stomach, and upper duodenum, while the midgut forms the lower duodenum, jejunum, ileum, cecum, appendix, ascending colon, and right half of the transverse colon.
The hindgut develops into the distal third of the transverse colon, descending colon, sigmoid colon, rectum, and upper portion of the anal canal.
The development of the digestive system can be affected by various anomalies, leading to conditions such as atresia, stenosis, inversion, and malpositioning of organs.
The digestive tube's various segments share a common organizational scheme, with minor local variations. Notably, development initiates in the upper regions before progressing to the lower ones. Initially, an epithelial tube, enveloped by splanchnic mesoderm, emerges. The mucosa undergoes rapid development, outpacing the other tunics, leading to the creation of folds. The mesenchymal covering undergoes differentiation, giving rise to the lamina propria, submucosa, muscular, and adventitious layers. Smooth muscle penetration into the mucosa results in the formation of the muscularis mucosae, causing a separation between the mucosa and submucosa. The circular muscle layer emerges between weeks six and ten, followed by the development of the one with longitudinal fibers between weeks ten and fourteen.
During the longitudinal folding of the embryo, the ectoderm at the cranial end forms a diverticulum, leading to the development of the oral cavity. Simultaneously, at the caudal pole, another diverticulum, known as the rectal cavity, takes shape.
The endoderm contributes to the epithelium of the digestive tube and the parenchyma of its accessory glands. In contrast, the mesoderm is responsible for forming the connective tissue, muscular components, and peritoneal structures. The foundation of the digestive tube and its accessory glands' differentiation lies in the reciprocal interactions between the endoderm (intestinal epithelium) and the surrounding mesoderm.
The divisions of the digestive tube are organized following the folding of the embryo. During this process, a portion of the yolk sac covered with endoderm is incorporated into the embryo's body, forming the primitive intestine. However, two other portions of this cavity, the yolk sac and the allantois, remain outside the embryo. The primitive intestine forms a tube in the cephalic and caudal portions of the embryo, with a blind sac at the end. The segments of this blind sac are called the proenteron and metenteron. The middle portion, known as the mesenteron, remains temporarily connected to the yolk sac through the vitelline duct or omphalo-enteric canal.
The development of the primitive intestine and its derived structures can be analyzed in segments. The pharyngeal intestine, or pharynx, extends from the level of the buccopharyngeal membrane to the tracheobronchial diverticulum. The anterior intestine, known as the proenteron, is located caudal to the pharyngeal intestine and extends to the hepatic primordium. It is vascularized by branches of the celiac artery. The middle intestine, called the mesenteron, starts from the hepatic primordium and continues to the area corresponding to the right two-thirds and left one-third junction of the transverse colon in adults. It is connected to the yolk sac by the vitelline duct and is vascularized by branches of the superior mesenteric artery. The posterior intestine, known as the metenteron, is located between the left third of the transverse colon and the cloacal membrane. It is vascularized by branches from the inferior mesenteric artery.
The endoderm is responsible for forming the epithelium that lines the interior (mucosa) of the digestive tube. It also gives rise to specific cells of the glands, such as hepatocytes and the exocrine and endocrine cells of the pancreas. On the other hand, the glandular stroma (connective tissue) and the muscular, connective, and peritoneal components of the intestinal wall are formed from the splanchnic mesoderm.
Mesenteries are double layers of peritoneum that anchor segments of the digestive tube and its derived structures to the dorsal and ventral walls of the trunk. They connect organs to the trunk wall and can be intraperitoneal or retroperitoneal. Peritoneal ligaments, which are also double layers of peritoneum, serve as pathways for blood vessels, nerves, and lymphatic vessels to travel to abdominal viscera.
Initially, the proenteron, mesenteron, and metenteron have a large contact surface with the mesenchyme of the posterior abdominal wall. However, by the end of the fifth week, a bridge of connective tissue narrows, and the caudal portion of the proenteron, mesenteron, and a significant part of the metenteron are suspended from the abdominal wall through the dorsal mesentery. Different regions of the digestive tube have specific mesenteries, such as the dorsal mesogastrium or greater omentum in the stomach region, the mesoduodenum in the duodenum, and the dorsal mesocolon in the region of the colon. The proper mesentery is formed by the dorsal mesentery corresponding to the jejunal and ileal loops.
On the other hand, the ventral mesentery is only present at the terminal segment of the esophagus, at the stomach, and in the upper part of the duodenum. It is derived from the transverse septum and is divided into two components by the development of the liver in the mesenchyme of the transverse septum. These components are the lesser omentum, which extends from the lower esophagus, stomach, and upper duodenum to the level of the liver, and the falciform ligament, which extends from the liver to the ventral wall of the trunk.
The Foregut, also known as the Anterior Intestine, is comprised of the branchial arches with branchial pouches at its cranial end. Around day 24, the pharyngeal membrane, consisting of ectodermal and endodermal layers, begins to disappear, allowing communication between the anterior primitive intestine and the amniotic cavity. The lower parts of the respiratory organs, followed by the esophagus, stomach, upper duodenum, liver, bile ducts, and pancreas, develop from the anterior intestine.
Specifically, the esophagus, stomach, proximal segment of the duodenum near the opening of the bile duct, tracheal buds, and lungs are formed from the foregut. On the other hand, the liver, pancreas, and biliary apparatus develop as evaginations of the endodermal epithelium corresponding to the upper portion of the duodenum. Within the upper portion of the anterior intestine, a septum known as the tracheo-esophageal septum divides it into the posteriorly located esophagus and the anteriorly located tracheal and lung buds. Any abnormalities in this septum can result in abnormal communication between the trachea and esophagus.
The development of the esophagus begins in the fourth week of gestation. At this stage, the respiratory diverticulum, also known as the pulmonary bud, emerges on the ventral wall of the anterior intestine, near the pharyngeal portion. As development progresses, the tracheo-esophageal septum separates the diverticulum from the dorsal portion of the proenteron. This division results in the formation of a ventral portion called the respiratory primordium and a dorsal portion known as the esophagus.
Initially, the esophagus is short but rapidly elongates during the growth of the neck and the descent of the heart and lungs, particularly through the growth of the cranial body. The epithelium of the esophagus transitions from a single layer to a double layer after the fourth week. In the second month, the epithelial cells multiply exponentially. By the third month, the epithelium becomes multi-layered and temporarily forms ciliated epithelium. Towards the end of pregnancy, the esophagus undergoes recanalization, increasing the lumen size, and the ciliated epithelium is replaced by a multi-layered flat epithelium. Additionally, esophageal glands develop in the surrounding mesenchyme.
During the second month of development, the mesenchyme surrounding the epithelial layer thickens and develops. The circular muscle appears first, followed by the longitudinal muscle in the second stage. The striated muscle in the upper third of the esophagus is derived from the last branchial arches and is innervated by the vagus nerve, the smooth muscle is derived from the splanchnopleura.
The muscular layer of the esophagus consists of striated muscle fibers in the upper two-thirds, which are innervated by the vagus nerve. In the lower third, the muscular layer consists of smooth muscle fibers that are innervated by the splanchnic plexus.
The development of the stomach begins in the fourth week of embryonic development in a 4mm embryo. Initially, it is a dilation of the anterior intestine with one cranial end and one caudal end. However, as the embryo grows, various factors cause changes in the appearance and position of the stomach.
One significant change is a 90° clockwise rotation of the stomach, known as dextrorotation. This rotation causes the left gastric surface to become anterior and the right surface to become posterior. As a result, the left vagus nerve, which originally innervated the left side of the stomach, now innervates the anterior gastric wall, while the right vagus nerve innervates the posterior gastric wall. During this rotation, the posterior wall of the stomach grows faster than the anterior wall, leading to the formation of the greater and lesser gastric curvatures.
Additionally, the cephalic and caudal segments of the stomach, initially located on the midline, undergo rotation around the anteroposterior axis. The caudal (pyloric) portion moves to the right and ascends, while the cephalic (cardia) portion moves to the left and slightly descends. This repositioning results in the final orientation of the gastric axis from the top-left to the bottom-right.
The stomach is attached to the dorsal wall of the trunk through the dorsal mesogastrium and to the ventral wall of the body through the ventral mesogastrium. The rotation and uneven growth of the stomach cause changes in the position of these mesenteries. The dorsal mesogastrium is pulled to the left, creating a space called the omental bursa (lesser peritoneal cavity) behind the stomach. Simultaneously, the ventral mesogastrium is pulled to the right due to this rotation.
During the development of the stomach, the primordium of the spleen emerges as a proliferation of mesoderm between the two layers of the dorsal mesogastrium. As a result of this rotation, the dorsal mesogastrium elongates and the portion between the spleen and the dorsal midline shifts to the left, merging with the peritoneum of the posterior abdominal wall. The posterior layer of the dorsal mesogastrium and the peritoneum along this fusion line degenerate.
The spleen remains in the peritoneal cavity and connects with the trunk wall near the left kidney through the splenorenal ligament, as well as with the stomach through the gastrosplenic ligament. This elongation and fusion of the dorsal mesogastrium also leads to the final positioning of the pancreas. Initially developing in the dorsal mesoduodenum, the pancreatic tail extends into the dorsal mesogastrium and becomes attached to it. After the degeneration of the posterior layer of the dorsal mesogastrium and the peritoneum of the posterior wall in the fusion line region, the pancreatic tail becomes retroperitoneal, covered by peritoneum only on its anterior surface.
The rotation of the stomach around the anteroposterior axis causes the dorsal mesogastrium to extend caudally and form a double-walled sac known as the greater omentum. This sac extends over the transverse colon and the loops of the small intestine, acting as a partition. The two layers of the greater omentum fuse to form a single cover suspended at the greater curvature of the stomach, while the posterior layer of the greater omentum fuses with the mesentery of the transverse colon.
The ventral mesogastrium, derived from the mesoderm of the transverse septum, gives rise to the lesser omentum and the falciform ligament. As the hepatic cords develop in the septum, it thins and forms the peritoneum of the liver. The falciform ligament connects the liver to the ventral wall of the trunk, while the lesser omentum extends from the stomach and upper duodenum to the liver. The free edge of the falciform ligament contains the umbilical vein, which obliterates after birth and forms the round ligament of the liver. The free edge of the lesser omentum, known as the hepatoduodenal ligament, contains the bile duct, portal vein, and hepatic artery. This free edge also forms the upper margin of Winslow's foramen, connecting the omental bursa (lesser peritoneal cavity) to the rest of the peritoneal cavity (greater peritoneal cavity).
The final position of the stomach allows for significant functional possibilities. It enables passive dilation of the stomach at the greater curvature during filling and facilitates active contraction movements during evacuation.
The development of the omental bursa begins in the fourth week with the appearance of spaces between mesenchymal cells in the dorsal mesogastrium. These spaces converge into a larger space, which forms the primordium of the omental bursa. The cavity extends transversely to the left and behind the stomach. A cranial extension, initially called the infracardiac bursa, is stopped by the development of the diaphragm muscle. Remnants of this infracardiac bursa can sometimes be found in adults. The lower portion of the cranial extension is known as the superior recess of the omental bursa.
As the stomach develops and rotates clockwise, the dorsal mesogastrium unfolds to the left, leading to a deepening of the omental bursa. This deepening forms the inferior recess, which gradually becomes a true sac that covers the transverse colon and small intestine in the last fetal months. After birth, the two layers of the sac fuse together, forming the greater omentum. Adipose tissue deposits in the greater omentum throughout life. The fusion of the two layers of the sac makes the inferior recess much smaller in adults.
Over time, the greater omentum merges with the posterior wall of the transverse mesocolon.
The omental bursa remains connected to the abdominal cavity through a slit-shaped opening called the epiploic foramen. This opening is located beneath the free edge of the hepatoduodenal ligament, which is part of the lesser omentum.
The development of the spleen begins in the fourth week and is closely linked to the mesenteries. The splenic primordia forms within the mesenchyme, specifically from the two layers of the dorsal mesogastrium, in front of the dorsal pancreatic bud. As it grows, the connection between the spleen and the dorsal mesogastrium becomes smaller, remaining only at the splenic hilum. The spleen maintains its position on the posterior abdominal wall and is attached to it by the lienorenal ligament. It is also connected to the stomach by the gastrolienal ligament. The splenic artery, which supplies blood to the spleen, originates from the celiac trunk located in the dorsal mesogastrium.
The definitive shape of the spleen is established in the third fetal month. In the fourth month, reticular tissue forms within the spleen, with lymphocytes appearing in the meshes. By the sixth month, these lymphocytes organize into white pulp nodules. During the third and fourth months of development, the mesenchyme from the dorsal mesogastrium forms the reticular connective tissue of the spleen, creating the splenic capsule and trabeculae inside the spleen. Around the beginning of the seventh month, the spleen begins to have hematopoietic function. The differentiation of white and red pulp in the spleen occurs only after birth.
The development of the duodenum begins with the formation of a bent segment of the intestine after the pylorus. This segment outlines the future duodenum and is included in the transverse septum.
The duodenum is formed from the terminal portion of the anterior intestine and the cephalic portion of the middle intestine. The junction between these components of the duodenum occurs just below the origin of the hepatic bud. The cranial half of the duodenum is supplied by branches from the celiac trunk, while the caudal half is supplied by the superior mesenteric artery. As the stomach rotates, the duodenum also rotates to the right, taking on a C-shaped loop shape.
During this rotation, the duodenum moves from its initial midline position to the left half of the abdominal cavity. The pancreatic head grows rapidly and comes into contact with the dorsal wall of the trunk. The right surface of the dorsal mesoduodenum fuses with the adjacent peritoneum. Eventually, both layers disappear, and the duodenum and pancreatic head become fixed in a retroperitoneal position.
The dorsal mesoduodenum completely disappears, except for a small segment of the duodenum (duodenal bulb) near the gastric pylorus, which retains its mesentery and remains intraperitoneal. The duodenal lumen is temporarily closed in the second month but reopens soon after. Duodenal glands appear in the fourth month as part of the regression process of epithelial cells. Kerckring's circular folds develop in the sixth month.
Since the proenteron is irrigated by the celiac artery and the mesenteron is irrigated by the superior mesenteric artery, the duodenum receives blood supply from both arteries.
The development of the liver and gallbladder begins with the formation of a hepatic territory at the lower end of the anterior intestine. In the third week, a hepatic groove appears, with a cranial portion that will become the liver and a caudal segment that will give rise to the gallbladder and bile ducts.
During the middle of the third week, the hepatic primordium emerges as an evagination of the endodermal epithelium. This hepatic diverticulum consists of rapidly proliferating cells that penetrate the transverse septum, separating the pericardial cavity from the yolk sac pedicle. As the hepatic cells penetrate the septum, the connection between the hepatic diverticulum and the anterior intestine narrows, forming the bile duct. A ventral evagination also appears, leading to the development of the gallbladder and cystic duct.
Bile secretion begins in the fourth month, flowing through the bile ducts into the intestine. The hepatic cords intertwine with the vitelline and umbilical veins, forming hepatic sinusoids. Hepatocytes and cells lining the inner surface of the bile ducts differentiate from the hepatic cords, while Kupffer cells, hematopoietic cells, and connective tissue cells derive from the mesoderm of the transverse septum.
The transverse septum fills with hepatic cells, becoming prominent in the abdominal cavity. The mesoderm between the liver and the proenteron forms the lesser omentum, while the mesoderm between the liver and the anterior abdominal wall forms the falciform ligament. This structure, known as the ventral mesentery, attaches the proenteron to the anterior abdominal wall.
The surface of the liver, except for the cranial surface, differentiates into visceral peritoneum. The compact mesoderm in a portion of the septum forms the central tendon of the diaphragm. The surface of the liver in contact with the future diaphragm remains uncovered by peritoneum, known as the bare area of the liver.
By week 10, the liver represents approximately 10% of the total body weight due to the large number of sinusoids and its hematopoietic function. However, this hematopoietic activity gradually diminishes during the last two months of intrauterine life, and at birth, only small islands of hematopoietic tissue remain. The liver's weight at birth is only 5% of the total body weight.
Around week 12, hepatic cells begin to secrete bile. As the gallbladder and cystic duct develop, the cystic duct joins the hepatic duct to form the common bile duct, allowing bile to reach the gastrointestinal tract. The bile duct gradually moves from an anterior to a posterior position due to changes in the position of the duodenum.
The development of the pancreas begins in the fifth week of vertebrate development, except for bony fishes. It originates from two buds, a larger dorsal bud and a smaller ventral bud, both derived from the endodermal layer of the duodenum. The dorsal bud is located in the dorsal mesentery, while the ventral bud is close to the bile duct. As the duodenum rotates, the ventral bud moves dorsally along with the bile duct. The ventral bud is situated inferior and posterior to the dorsal bud, and their parenchyma and ductal systems merge. The uncinate process and lower portion of the pancreatic head develop from the ventral bud, while the rest of the gland forms from the dorsal bud.
During the seventh week, the two pancreatic primordia merge, causing changes in the positioning and drainage pattern of the biliary and pancreatic ducts into the duodenum. The unified pancreatic primordia are located in the primitive dorsal mesentery, covered on both sides by peritoneum. Due to changes in the positions of the stomach and duodenum, the pancreas becomes parallel to the posterior abdominal wall, even fusing with it. This transition from an initially intraperitoneal organ to a secondary retroperitoneal organ occurs.
The body and tail of the pancreas, as well as the cranial portion of the head, originate from the dorsal pancreatic primordium. The caudal part of the pancreatic head originates from the ventral primordium.
The main pancreatic duct (Wirsung's duct) is formed from the distal segment of the dorsal pancreatic duct and the entire ventral pancreatic duct. The proximal portion of the dorsal pancreatic duct either disappears or persists as a narrow canal called the accessory pancreatic duct (Santorini's duct). The main pancreatic duct and the bile duct enter the duodenum at the major duodenal papilla. If the accessory duct is present, it enters the duodenum at the minor duodenal papilla. In approximately 10% of cases, the ductal systems of the two buds do not fuse, resulting in the persistence of the initial double ductal system.
Pancreatic islets (of Langerhans) develop from the pancreatic parenchyma in the third month of fetal life. They are distributed diffusely throughout the entire pancreas and have recently been discovered to have origins in neural crest cells. Insulin secretion begins around the fifth month. Parenchymal cells also give rise to cells secreting glucagon and somatostatin. The pancreatic connective tissue is formed by the splanchnic mesoderm surrounding the pancreatic buds. Insulin production starts as early as the fifth fetal month.
The Mesenteron, also known as the midgut, is formed from the middle portion of the primitive intestine. This includes the lower part of the duodenum, jejunum, ileum, cecum, vermiform appendix, ascending colon, and the right half of the transverse colon. Vascularization of these intestinal portions is provided by branches from the superior mesenteric artery. The development of the midgut is influenced by significant shape and positional changes, with the development of the umbilical loop and physiological umbilical detachment being key features.
By the end of the fifth week, the midgut is attached to the dorsal abdominal wall by a short mesentery and is directly connected to the yolk sac through the vitelline duct or vitelline pedicle. In adults, the midgut begins just below the entry point of the bile duct into the duodenum and ends at the junction between the proximal two-thirds and the distal third of the transverse colon. The superior mesenteric artery supplies blood to the midgut.
During the first month, a 90-degree counterclockwise rotation of the umbilical loop begins. This results in the two arms of the loop being positioned in a transverse plane, with the cranial arm on the right and the caudal arm on the left. In the following months, the small intestine undergoes growth and development.
The right arm of the umbilical loop lengthens, forming multiple loops, and eventually, the left arm also develops intestinal loops. At this stage of development growing, there is limited space in the trunk cavity. As a result, the intestine is pushed forward into the coelomic umbilical cord.
The development of the midgut is characterized by the rapid elongation of the intestinal tube and its mesentery, with the formation of the primary intestinal loop. This loop communicates with the yolk sac through a narrow vitelline duct. The distal portion of the duodenum, jejunum, and part of the ileum are formed by the cephalic segment of the loop, while the caudal segment forms the lower portion of the ileum, cecum, appendix, ascending colon, and the proximal two-thirds of the transverse colon.
During the intestinal rotation (of 270 - 300 degrees), the cecum initially migrates superiorly and then from the left to the right below the liver. It then extends inward, reaching the right iliac fossa. These position changes are accompanied by the formation of flexures that give the large intestine its definitive segments.
At the distal end of the cecum, the vermiform appendix is formed. Only in the seventh to eighth month is there a clear delineation between the cecum and the vermiform appendix, and the appendix can also form loops. Around the tenth week, the intestinal loops retract posteriorly into the abdominal cavity.
During the development of the primary intestinal loop, the cephalic segment undergoes rapid elongation. This growth is accompanied by the rapid growth and development of the liver. As a result, the abdominal cavity becomes temporarily too small to accommodate the intestinal loops. Consequently, in the sixth week of development, the intestinal loops enter the extraembryonic cavity within the umbilical cord. This phenomenon is known as physiological umbilical hernia.
During the development of the primary intestinal loop, the cephalic segment undergoes rapid elongation. This growth is accompanied by the rapid growth and development of the liver. As a result, the abdominal cavity becomes temporarily too small to accommodate the intestinal loops. Consequently, in the sixth week of development, the intestinal loops enter the extraembryonic cavity within the umbilical cord. This phenomenon is known as physiological umbilical hernia.
Rotation of the Midgut occurs as the primary intestinal loop lengthens. It rotates counterclockwise around the axis formed by the superior mesenteric artery. The rotation is 270° when viewed from the anterior perspective. During rotation, the small intestine continues to elongate, forming multiple loops with the jejunum and ileum. The large intestine elongates significantly but does not participate in the twisting process. Rotation happens during both physiological herniation (approximately 90°) and when the intestinal loops return to the abdominal cavity (the remaining 180°).
In the development of the primary intestinal loop, the cephalic segment experiences rapid elongation. This growth is accompanied by the rapid growth and development of the liver. Consequently, the abdominal cavity becomes temporarily too small to accommodate the intestinal loops. This leads to the intestinal loops entering the extraembryonic cavity within the umbilical cord during the sixth week of development. This phenomenon is referred to as physiological umbilical hernia.
During the 10th week, the herniated intestinal loops retract back into the abdominal cavity. This retraction is caused by the regression of the mesonephric kidney, a decrease in the liver's growth rate, and the expansion of the abdominal cavity.
The first segment to return to the abdominal cavity is the proximal portion of the jejunum, which is located on the left side. Subsequently, the remaining loops progressively arrange themselves towards the right side. The cecal primordium, which appears as a small conical dilation of the caudal segment of the primary intestinal loop around the sixth week, is the last portion of the intestine to retract. Initially, it is positioned in the upper right abdominal quadrant, just below the right hepatic lobe. It then descends into the right iliac fossa, causing the ascending colon and hepatic flexure to move to the right side of the abdominal cavity. The distal end of the cecal primordium develops into a narrow diverticulum known as the appendix. As the colon descends, the appendix will ultimately be positioned posterior to the cecum or colon, in retrocecal and retrocolic positions, respectively.
The mesenteries of the intestinal loops are bands that anchor internal organs, containing nerves, blood vessels, and lymphatic vessels for the organs they fix. They consist of two serous layers with a thin sheet of connective tissue between them, derived from the peritoneum.
During the rotation and formation of intestinal loops, the mesentery of the primary intestinal loop (the actual mesentery) undergoes significant transformations. The dorsal mesentery twists around the origin of the superior mesenteric artery as the caudal segment of the primary loop moves to the right side of the abdominal cavity. The mesenteries of the ascending and descending portions of the colon come into close contact with the peritoneum of the posterior abdominal wall when they reach their final positions. The ascending and descending colons remain fixed in a retroperitoneal position after the fusion of these serous layers, while the appendix, lower end of the cecum, and sigmoid colon retain their own free mesentery.
The transverse mesocolon fuses with the posterior wall of the greater omentum, maintaining its mobility. The attachment line of the transverse mesocolon extends from the hepatic flexure of the ascending colon to the splenic flexure of the descending colon. In areas where the mesenteries begin and end their attachments, peritoneal recesses called peritoneal niches are formed where the fusion of the mesentery layers remained incomplete. Examples of these recesses include the superior and inferior duodenal recesses around the duodenojejunal flexure, the superior and inferior ileocecal recesses in the cecal region, and the intersigmoid recess at the level of the sigmoid mesocolon.
The mesentery of the jejuno-ileal loops initially continues with the mesentery of the ascending colon. However, when the mesentery of the ascending mesocolon fuses with the posterior abdominal wall, the mesentery of the jejuno-ileal loops acquires a new attachment line that extends from the area where the duodenum becomes intraperitoneal to the ileocecal junction.
The ventral mesogastrium and ventral mesoduodenum form the peritoneal covering of the liver beneath the diaphragm. The extensive connection with the transverse septum is greatly reduced due to the growth and enlargement of the liver, resulting in a small area known as the bare area of the liver. This bare area represents a direct attachment of the liver to the diaphragm, delimited by the coronary ligament of the liver. The rotation of the stomach and intestines during development leads to the distinctive final appearance of shape and positioning, with the liver stretched towards the right lateral direction and the pancreas towards the left lateral direction.
The primitive dorsal mesentery subdivides into the dorsal mesoesophagus, dorsal mesogastrium, dorsal mesoduodenum, and common dorsal mesentery. The common dorsal mesentery fixes the jejunum, ileum, colon, and rectum to the posterior abdominal wall.
The hindgut, also known as the posterior intestine, extends from the midgut to the level of the cloacal membrane. It is supplied by the inferior mesenteric artery. The lower end of the hindgut flares out and forms part of the cloaca, where the primitive renal ducts drain on both sides.
From the hindgut, the distal third of the transverse colon, descending colon, sigmoid colon, rectum, and upper portion of the anal canal develop. The endoderm of the posterior intestine forms the mucosa of the urinary bladder and urethra. The terminal portion of the hindgut enters the posterior region of the cloaca, which represents the primitive ano-rectal canal. The allantois enters the anterior portion of the cloaca, representing the primitive uro-genital sinus. The cloaca is a cavity lined with endoderm and covered ventrally by the surface ectoderm. The boundary between the ectoderm and endoderm forms the cloacal membrane. The allantois is separated from the hindgut by a layer of mesoderm, forming the uro-rectal septum derived from the merging portions of the mesoderm covering the yolk sac and surrounding the allantois.
During embryonic development, the caudal folding continues, and the tip of the uro-rectal septum approaches the cloacal membrane. However, these two structures do not come into contact. At the end of the seventh week, the cloacal membrane perforates, forming the anal opening of the posterior intestine and the ventral opening of the urogenital sinus. The uro-rectal septum forms the perineal body between these openings.
The ectoderm proliferates and closes the caudal region of the anal canal. In the ninth week, this region becomes repermeabilized. The inferior rectal arteries, branches of the internal pudendal arteries, irrigate the caudal segment of the anal canal, which has an ectodermal origin. The cranial segment of the anal canal is irrigated by the superior rectal artery, a continuation of the inferior mesenteric artery, and has an endodermal origin.
The pectinate (dentate) line marks the junction between the endodermal and ectodermal regions of the anal canal. It separates the columnar epithelium of the rectum from the stratified squamous epithelium of the anal canal. The pectinate line is located just below the anal columns (Morgagni). Abnormalities in the dimensions of the posterior cloacal region can cause the anterior displacement of the anal opening, resulting in the formation of recto-vaginal and recto-urethral fistulas and atresias.
There are several anomalies that can occur during the development of the esophagus.
One such anomaly is atresia or congenital obliteration. This happens when the epithelial plug fails to be resorbed, often accompanied by tracheoesophageal fistulas. As a result, amniotic fluid cannot enter the fetal digestive tract, leading to the development of hydramnion.
Another congenital anomaly is esophageal stenosis. This condition causes ingested food to be expelled through vomiting.
The short esophagus is another developmental anomaly. In this case, the esophagus does not reach below the diaphragm and is often accompanied by a partially thoracic stomach.
Esophageal diverticula are also a result of developmental anomalies. They occur when there are abnormalities in the separation of the pulmonary bud from the esophagus.
There are several anomalies that can occur in the development of the stomach. One such anomaly is the congenital bilocular stomach, which is caused by an uneven development of the circular muscular layer.
Another anomaly is gastroschisis, which is characterized by the absence of the abdominal wall, typically on the right side in males. This condition results in the viscera being exposed to amniotic fluid.
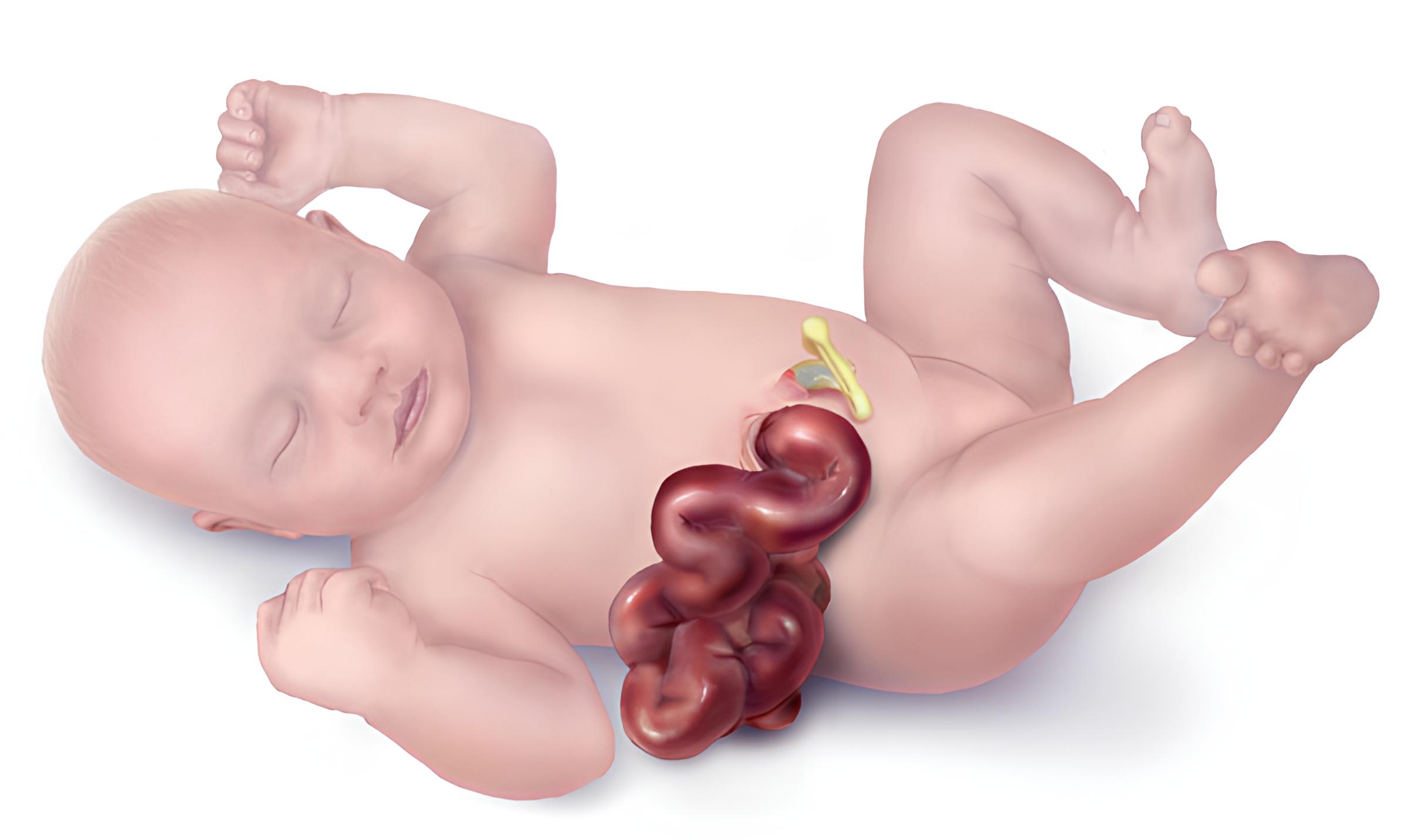
Stomach inversion is a condition that occurs when the viscera undergo a general inversion, known as situs inversus. In this case, the stomach also undergoes inversion.
Thoracic stomach is another anomaly that can occur. It is caused by a descent anomaly and a short esophagus, which leads to the cardia being located in the posterior mediastinum.
Lastly, there is congenital pyloric atresia or stenosis. Congenital pyloric stenosis is caused by an exaggerated development of the pyloric sphincter. This condition typically manifests after birth.
Observations of anomalies in the development of the spleen include incomplete fusion of primary lobes and the presence of accessory spleens.
There are two types of anomalies in the development of the duodenum: duodenal stenosis and duodenal atresia. Duodenal stenosis is a result of incomplete recanalization of the digestive tube and is commonly found in the horizontal and ascending segments of the duodenum. This condition is manifested in newborns through bilious vomiting. On the other hand, duodenal atresia occurs when the descending and horizontal segments of the duodenum are completely obliterated, specifically caudal to the outlet of the common bile duct.
The liver and gallbladder can experience anomalies in their development. These anomalies include lobulation and fissuration abnormalities, which can lead to the persistence of Riedel's lobe, an elongation of the right lobe, even after birth. Additionally, there may be the presence of accessory hepatic ducts, with the cystic duct opening into one of them. Temporary obliteration processes can also occur, resulting in atresia of the extrahepatic bile ducts. Another anomaly is the occurrence of a bifid or double gallbladder, where the bladder bud partially or completely divides.
The development of the pancreas can sometimes result in anomalies. One such anomaly is known as ring pancreas, which occurs when the ventral bud does not move and remains in front of the duodenum. In this case, the ventral bud, along with the dorsal bud, forms a ring around the duodenum. This condition can lead to duodenal stenosis or even complete obstruction. Ring pancreas is more common in males.
Another anomaly can occur when the pancreatic bud migrates with the hepatic bud. This can result in the formation of a nodule or pancreatic vesicle near the gallbladder.
In addition, masses of pancreatic tissue can be found in various locations such as the walls of the stomach, intestines, or in relation to the spleen. These masses are referred to as aberrant pancreas.
The development of the intestines can be affected by several anomalies. One such anomaly is Meckel's diverticulum, which is found on the antimesenteric border of the ileum. This diverticulum may contain gastric or pancreatic tissue and can cause ulcers and bleeding when secretions are released.
Another anomaly is the absence of abdominal wall formation, which leads to the eventration of the viscera. In this condition, the viscera are only covered by the amnion and peritoneum.
Displacement and coalescence processes can also result in various anomalies. If coalescence processes do not occur, the intestines remain free and are suspended by a free mesentery. This can lead to complications such as twisting, which can cause complete or incomplete volvulus or internal hernia.
The development of the digestive system involves the formation and differentiation of various structures, including the esophagus, stomach, liver, gallbladder, pancreas, and intestines.
Development begins with the emergence of an epithelial tube enveloped by splanchnic mesoderm. The mucosa undergoes rapid development, leading to the formation of folds, while the mesenchymal covering differentiates into various layers.
The digestive tube is divided into different segments, each with specific characteristics and functions. Mesenteries, double layers of peritoneum, anchor segments of the digestive tube to the trunk walls and allow for the passage of blood vessels and nerves.
The foregut gives rise to the esophagus, stomach, and upper duodenum, while the midgut forms the lower duodenum, jejunum, ileum, cecum, appendix, ascending colon, and right half of the transverse colon.
The hindgut develops into the distal third of the transverse colon, descending colon, sigmoid colon, rectum, and upper portion of the anal canal.
The development of the digestive system can be affected by various anomalies, leading to conditions such as atresia, stenosis, inversion, and malpositioning of organs.
Development of the Digestive System, digestive tube, splanchnic mesoderm, mucosa, folds, lamina propria, submucosa, muscular layer, adventitious layer, muscularis mucosae, circular muscle layer, longitudinal fibers, ectoderm, diverticulum, endoderm, connective tissue, peritoneal structures, mesenteries, peritoneal ligaments, blood vessels, nerves, lymphatic vessels, foregut, pharyngeal intestine, proenteron, mesenteron, metenteron, endoderm, glands, mesoderm, mesenteries, dorsal mesogastrium, ventral mesentery, Foregut, Anterior Intestine, esophagus, tracheobronchial diverticulum, stomach, duodenum, liver, bile ducts, pancreas, rotation, dextrorotation, vagus nerve, lesser omentum, falciform ligament, omental bursa, spleen, pancreatic tail, greater omentum, ventral mesentery, stomach rotation, duodenum, liver, gallbladder, hepatic primordium, bile duct, pancreatic bud, hepatic diverticulum, hepatic cords, hepatic sinusoids, pancreatic islets, duodenum, jejunum, ileum, cecum, vermiform appendix, ascending colon, transverse colon, Midgut, umbilical loop, physiological umbilical hernia, intestinal rotation, retraction of herniated loops, mesenteries, dorsal mesentery, ventral mesogastrium, bare area of the liver, gastrolienal ligament, hindgut, cloaca, anal canal, pectinate line, malformations, esophageal atresia, esophageal stenosis, short esophagus, esophageal diverticula, gastroschisis, stomach inversion, thoracic stomach, pyloric atresia, spleen anomalies, incomplete fusion, accessory spleens, duodenal stenosis, duodenal atresia, liver anomalies, lobulation abnormalities, accessory hepatic ducts, atresia of extrahepatic bile ducts, bifid gallbladder, pancreas anomalies, ring pancreas, pancreatic vesicle, aberrant pancreas, Meckel's diverticulum, eventration, twisting, volvulus, internal hernia.Embryonic Development and Anomalies of the Digestive Tube: An OverviewDevelopment of the Digestive System0000